The nigrostriatal pathway is one of the brain’s most critical neural circuits — a dopaminergic highway connecting the midbrain to the basal ganglia that makes smooth, coordinated, voluntary movement possible. Without it, the simple act of reaching for a glass of water, standing up from a chair, or beginning to walk would become effortful, rigid, and tremulous. Most people encounter this pathway’s name only when something goes wrong with it — most famously in Parkinson’s disease, where the progressive loss of dopamine-producing neurons in this circuit transforms movement into a daily challenge.
But the nigrostriatal pathway is far more than a clinical footnote. It is a masterpiece of neurological engineering: a finely balanced circuit of excitatory and inhibitory signals that acts as the brain’s gatekeeper for movement, continuously deciding which motor programs to allow and which to suppress. It integrates signals from the cortex, filters them through the complex circuitry of the basal ganglia, and returns a verdict to the thalamus and motor cortex about whether and how to move.
Understanding this pathway matters not only for neuroscientists and clinicians but for anyone interested in how the brain governs the body — and why conditions as different as Parkinson’s disease, Huntington’s disease, and the motor side effects of antipsychotic medications all converge on this same anatomical corridor. Knowing how dopamine functions within this system helps explain why its disruption is so devastating, why current treatments work the way they do, and what directions future research is pursuing.
This article explores the nigrostriatal pathway in depth: its anatomy, its key structures, the direct and indirect pathways through which it operates, its roles in motor control and beyond, the conditions that arise when it is damaged, and the therapeutic approaches that attempt to restore its function.
What Is the Nigrostriatal Pathway? A Clear Definition
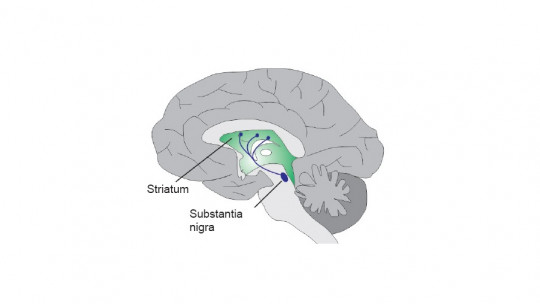
The nigrostriatal pathway is a dopaminergic neural tract that originates in the substantia nigra pars compacta (SNc) of the midbrain and projects to the striatum — specifically the caudate nucleus and putamen — which are the primary input structures of the basal ganglia. The name itself describes the anatomy: nigro from the substantia nigra, and striatal from the striatum.
It is one of four major dopaminergic pathways in the brain, alongside the mesolimbic, mesocortical, and tuberoinfundibular pathways. Of these four, the nigrostriatal pathway is the longest and carries the largest proportion of the brain’s total dopamine. Its principal function is motor control — regulating the initiation, coordination, and smooth execution of voluntary movement.
The pathway operates through dopamine’s action on two families of receptors in the striatum:
- D1 receptors — coupled to the direct pathway, through which dopamine facilitates movement by ultimately disinhibiting the thalamus
- D2 receptors — coupled to the indirect pathway, through which dopamine suppresses movement-inhibiting circuits, also ultimately promoting movement
This dual-receptor system allows dopamine to modulate both the accelerator and brake of the motor circuit simultaneously — a level of fine-tuned control that explains why even partial disruption of dopamine availability in this pathway produces such pronounced motor consequences.
| Feature | Detail |
|---|---|
| Origin | Substantia nigra pars compacta (SNc), midbrain |
| Destination | Striatum (caudate nucleus + putamen), basal ganglia |
| Neurotransmitter | Dopamine |
| Primary function | Motor control, movement initiation and coordination |
| Key receptors | D1 (direct pathway) and D2 (indirect pathway) |
| Associated disorder | Parkinson’s disease, drug-induced parkinsonism |
Key Brain Structures That Make Up the Nigrostriatal Pathway
To understand how this pathway functions, it helps to understand each of its key anatomical components — not as isolated structures but as participants in an integrated circuit.
The substantia nigra is a small but remarkably important region of the midbrain, located in the ventral mesencephalon. Its name means “black substance” in Latin, a reference to the dark pigmentation of its neurons caused by neuromelanin — a byproduct of dopamine synthesis. The substantia nigra is divided into two distinct zones:
- Substantia nigra pars compacta (SNc): the densely packed region that houses the dopamine-producing neurons that project to the striatum. This is the origin of the nigrostriatal pathway and the region most devastated in Parkinson’s disease
- Substantia nigra pars reticulata (SNr): the larger, more diffuse region that functions as a major output nucleus of the basal ganglia, projecting inhibitory GABAergic signals to the thalamus and brainstem
The striatum is the largest structure within the basal ganglia and the primary destination of nigrostriatal dopamine. It is composed of two principal structures:
- Caudate nucleus: a C-shaped structure involved in motor planning, cognitive aspects of movement, and procedural learning. The caudate is also connected to frontal cortical regions and plays a role in executive function and habit formation
- Putamen: closely linked to the caudate and primarily involved in the regulation of movement and motor learning. The putamen is the striatal region most directly involved in the nigrostriatal pathway’s moment-to-moment motor control
Together, the caudate and putamen are collectively called the neostriatum or simply the striatum. They receive input from virtually the entire cortex via the corticostriatal pathway, making them a remarkable convergence zone where cognitive, sensory, emotional, and motor information is integrated before being filtered through basal ganglia circuits.
Beyond the SNc and striatum, the full nigrostriatal circuit also involves the globus pallidus (internal and external segments), the subthalamic nucleus, and the thalamus — all of which participate in the direct and indirect pathways described in the next section.

The Direct and Indirect Pathways: How the Nigrostriatal Circuit Controls Movement
The nigrostriatal pathway does not control movement directly. Instead, it modulates two competing basal ganglia circuits — the direct pathway and the indirect pathway — that together act as a finely tuned gate for motor output. Understanding both is essential to understanding how dopamine loss produces movement disorders.
The basal ganglia maintain a default state of tonic inhibition of the thalamus and brainstem motor nuclei. Movement occurs when this inhibitory tone is selectively reduced — essentially lifting the brake — to allow the thalamus to excite the motor cortex and generate movement. The direct and indirect pathways represent two complementary mechanisms for achieving this.
The direct pathway promotes movement through the following sequence:
- The motor cortex sends excitatory glutamatergic signals to the striatum (caudate and putamen)
- Activated striatal neurons project inhibitory GABAergic signals to the globus pallidus internal (GPi) and substantia nigra pars reticulata (SNr)
- GPi/SNr activity is reduced — meaning their tonic inhibition of the thalamus is released
- The thalamus becomes active and sends excitatory signals to the motor cortex
- Movement is facilitated
Dopamine from the SNc acting on D1 receptors in the striatum enhances the activity of this direct pathway, amplifying the pro-movement signal.
The indirect pathway suppresses unwanted movements through a longer, more complex route:
- The cortex excites the striatum
- Activated striatal neurons inhibit the globus pallidus external (GPe)
- Reduced GPe activity disinhibits the subthalamic nucleus (STN)
- An active STN sends excitatory signals to GPi/SNr
- GPi/SNr strongly inhibits the thalamus, suppressing movement
Dopamine acting on D2 receptors in the striatum suppresses the indirect pathway — meaning it reduces the movement-inhibiting signal, again producing a net pro-movement effect. This is why dopamine is described as the molecule that keeps the motor gate open: it simultaneously amplifies the direct pathway and dampens the indirect one.
When dopamine is depleted, both effects are lost simultaneously: the direct pathway loses its amplification, and the indirect pathway loses its suppression. The result is an overactive inhibitory output from GPi/SNr, a chronically suppressed thalamus, and severely impaired ability to initiate and sustain voluntary movement — the clinical picture of Parkinson’s disease.

The Role of Dopamine: Why This Neurotransmitter Is So Central
Dopamine is the biochemical currency of the nigrostriatal pathway. Without it, the exquisitely balanced circuit of excitation and inhibition described above collapses into a state of pathological inhibition that makes movement effortful, slow, and imprecise.
The dopaminergic neurons of the substantia nigra pars compacta are among the most metabolically demanding neurons in the brain. They maintain an unusual autonomous pacemaker activity — firing at a low, steady rate even in the absence of synaptic input — which generates a tonic baseline of dopamine in the striatum. This tonic dopamine level serves as a kind of ready state for the motor system, maintaining the direct-indirect pathway balance in a configuration that allows rapid, smooth movement on demand.
When a movement is initiated, phasic bursts of dopamine are released on top of this tonic baseline, reinforcing the striatal circuits involved in that specific movement. This phasic signaling is thought to play a critical role not only in movement execution but in motor learning — the dopamine burst acts as a reinforcement signal that strengthens the neural circuits underlying successful movements over time. This is why the nigrostriatal pathway is essential not only for moving but for learning how to move skillfully.
The vulnerability of SNc dopaminergic neurons is one of the most important unsolved questions in neuroscience. Their high metabolic demands, their exposure to oxidative stress from dopamine metabolism, and their particular mitochondrial characteristics all contribute to making them disproportionately susceptible to the aging-related and pathological processes that underlie Parkinson’s disease.
Nigrostriatal Pathway and Parkinson’s Disease: What Goes Wrong
Parkinson’s disease is, at its neurochemical core, a disease of the nigrostriatal pathway. It develops when the dopaminergic neurons of the substantia nigra pars compacta undergo progressive degeneration — a process that unfolds slowly over years before the motor symptoms become clinically apparent. By the time a person receives a Parkinson’s diagnosis, they have typically already lost 50–70% of their SNc dopaminergic neurons and depleted striatal dopamine by a similar magnitude.
The cardinal motor symptoms of Parkinson’s disease map directly onto the functional consequences of nigrostriatal dopamine loss:
- Bradykinesia — slowness of movement, the most characteristic feature, reflecting the reduced thalamic activation that results from overactive GPi/SNr inhibition
- Resting tremor — the characteristic 4–6 Hz “pill-rolling” tremor that occurs at rest, thought to arise from oscillatory activity in the basal ganglia-thalamo-cortical circuit when normal dopaminergic modulation is absent
- Rigidity — increased muscle tone and resistance to passive movement, reflecting abnormal co-activation of agonist and antagonist muscle groups
- Postural instability — impaired ability to maintain upright posture and make rapid postural corrections, emerging typically in later stages
- Freezing of gait — the episodic inability to initiate or continue walking, one of the most disabling features of advanced Parkinson’s disease
The neuropathological signature of Parkinson’s disease is the Lewy body — an abnormal intracellular aggregate composed primarily of misfolded alpha-synuclein protein — first described by Friedrich Heinrich Lewy in 1912. Braak and colleagues subsequently proposed a staging model suggesting that alpha-synuclein pathology begins in the brainstem and olfactory bulb and ascends through the SNc and eventually to cortical regions, providing a potential explanation for the progression of both motor and non-motor symptoms over time.
The current standard of care for motor symptoms — levodopa, a dopamine precursor that crosses the blood-brain barrier and is converted to dopamine in the brain — was introduced in the late 1960s by George Cotzias and remains the most effective pharmacological treatment available. Its effectiveness is itself powerful evidence for the centrality of dopamine in the nigrostriatal pathway’s function.

Beyond Movement: Other Functions of the Nigrostriatal Circuit
While motor control is the nigrostriatal pathway’s primary and best-characterized function, research over the past two decades has revealed that its contributions extend meaningfully beyond movement regulation.
Procedural and habit learning depends significantly on the striatum and its dopaminergic modulation. Ann Graybiel’s influential research on basal ganglia function has demonstrated that the striatum is essential for the acquisition and storage of habitual behaviors — the automatic, chunked motor sequences that allow us to perform well-practiced actions (driving, typing, playing an instrument) without conscious deliberation. Dopamine in the nigrostriatal pathway appears to function as a reinforcement signal that consolidates successful motor patterns into automatic habits.
The caudate nucleus in particular has been implicated in higher-order aspects of movement including motor planning, the selection among competing movement options, and the integration of cognitive goals with motor execution. Patients with Parkinson’s disease show deficits not only in movement execution but in motor planning and sequence learning, reflecting the role of caudate dopamine in these processes.
There is also growing evidence that the nigrostriatal pathway contributes to reward processing and motivational aspects of movement — specifically, the motivational cost-benefit calculation of whether a given physical action is worth the effort required. When dopamine is depleted, movement becomes not only harder to execute but harder to motivate — a phenomenon that may contribute to the apathy and reduced physical activity commonly observed in Parkinson’s disease even in early stages.
Drug-Induced Disruption: Antipsychotics and Extrapyramidal Symptoms
The nigrostriatal pathway is the anatomical explanation for one of the most significant problems in psychopharmacology: extrapyramidal symptoms (EPS) caused by antipsychotic medications.
Antipsychotic drugs — both first-generation (typical) and, to a lesser degree, second-generation (atypical) antipsychotics — work primarily by blocking D2 dopamine receptors. This D2 blockade in the mesolimbic pathway reduces the dopaminergic hyperactivity that is thought to underlie psychotic symptoms. However, D2 receptors are not exclusive to the mesolimbic pathway. They are abundantly expressed in the striatum, where they are essential to the indirect pathway’s proper function.
When antipsychotics block D2 receptors in the striatum, they effectively replicate the functional consequences of dopamine depletion in the nigrostriatal pathway — producing a drug-induced parkinsonism that can be clinically indistinguishable from idiopathic Parkinson’s disease. The resulting extrapyramidal symptoms include:
- Pseudoparkinsonism: tremor, rigidity, and bradykinesia identical to Parkinson’s motor features
- Akathisia: a deeply uncomfortable sensation of inner restlessness and compulsion to move, often described by patients as more distressing than the original psychiatric symptoms being treated
- Acute dystonia: sudden, involuntary muscle contractions causing abnormal postures, particularly of the neck, face, and limbs
- Tardive dyskinesia: a potentially irreversible syndrome of involuntary, repetitive movements — typically involving the mouth, tongue, and face — that emerges with prolonged antipsychotic use, thought to result from D2 receptor upregulation in the striatum
The development of atypical antipsychotics — including clozapine, quetiapine, and others — was motivated in significant part by the desire to achieve antipsychotic efficacy while minimizing nigrostriatal D2 blockade, thereby reducing EPS. Understanding the nigrostriatal pathway is therefore not merely a neuroscience matter but a directly clinically relevant one that shapes how psychiatric medications are designed and monitored.

The Nigrostriatal Pathway in Huntington’s Disease: The Opposite Disruption
Where Parkinson’s disease results from the loss of nigrostriatal dopamine input to the striatum, Huntington’s disease represents a fundamentally different disruption of the same circuit — one that produces the opposite motor phenotype.
Huntington’s disease is caused by an autosomal dominant mutation in the HTT gene, producing an abnormally long polyglutamine repeat in the huntingtin protein. The striatum — particularly the caudate nucleus and putamen — undergoes progressive neurodegeneration, with the medium spiny neurons of the indirect pathway being selectively vulnerable in the early stages. Because the indirect pathway normally suppresses unwanted movements, its early degeneration reduces the brain’s capacity to inhibit movement, producing the characteristic chorea — involuntary, irregular, dance-like movements — that gives the disease its name (from the Greek choreia, meaning dance).
As Huntington’s disease progresses and the direct pathway is also damaged, the hyperkinetic chorea can give way to a more rigid, hypokinetic syndrome resembling Parkinson’s disease — a progression that dramatically illustrates how the balance between the direct and indirect pathways determines the overall motor phenotype.
Therapeutic Approaches Targeting the Nigrostriatal Pathway
The clinical significance of the nigrostriatal pathway has driven decades of therapeutic innovation. Current and emerging approaches span pharmacological, surgical, and experimental biological strategies.
Pharmacological approaches: Levodopa (L-DOPA), combined with carbidopa to prevent peripheral conversion, remains the gold standard for Parkinson’s motor symptom management. Dopamine agonists (pramipexole, ropinirole) directly stimulate dopamine receptors and are often used in early-stage disease or as adjuncts to levodopa. MAO-B inhibitors (selegiline, rasagiline) reduce dopamine breakdown. COMT inhibitors (entacapone) extend levodopa’s duration of action. Each of these strategies ultimately aims at the same goal: restoring adequate dopaminergic signaling in the striatum.
Deep brain stimulation (DBS) has transformed the management of advanced Parkinson’s disease since its development by Alim-Louis Benabid and colleagues in the 1980s and 1990s. By implanting electrodes in the subthalamic nucleus or GPi and delivering continuous high-frequency electrical stimulation, DBS effectively disrupts the pathologically overactive output of these structures — functionally mimicking the effect of dopamine in reducing GPi inhibition of the thalamus. DBS does not replace dopamine but compensates for its absence through a different mechanism in the same circuit.
Cell and gene therapies represent the frontier of nigrostriatal pathway research. Approaches under investigation include the transplantation of dopamine-producing neurons derived from human embryonic stem cells or induced pluripotent stem cells (iPSCs), viral vector-mediated delivery of neurotrophic factors (particularly GDNF, glial cell line-derived neurotrophic factor) to protect remaining SNc neurons, and gene editing approaches targeting alpha-synuclein aggregation. While none of these has yet achieved clinical translation at scale, several are in active clinical trials and represent the most promising paths toward disease-modifying rather than merely symptomatic treatment.
FAQs about the Nigrostriatal Pathway
What is the nigrostriatal pathway and what does it do?
The nigrostriatal pathway is a dopaminergic neural tract that connects the substantia nigra pars compacta in the midbrain to the striatum (caudate nucleus and putamen) — the main input structures of the basal ganglia. Its primary function is motor control: it modulates the direct and indirect pathways of the basal ganglia circuit, regulating the initiation, coordination, and smooth execution of voluntary movement. By acting on D1 and D2 receptors in the striatum, dopamine from the SNc simultaneously amplifies the movement-promoting direct pathway and suppresses the movement-inhibiting indirect pathway, maintaining the basal ganglia’s motor gate in a state that allows fluid, purposeful movement on demand.
What happens when the nigrostriatal pathway is damaged?
When the nigrostriatal pathway is damaged — specifically when dopaminergic neurons in the substantia nigra pars compacta degenerate — the loss of dopamine input to the striatum disrupts the balance between the direct and indirect pathways. The indirect pathway becomes overactive, producing excessive inhibition of the thalamus and motor cortex. The clinical result is the movement disorder characteristic of Parkinson’s disease: bradykinesia (slowed movement), resting tremor, muscular rigidity, postural instability, and difficulty initiating movement. Pharmacological disruption of D2 receptors in this pathway — by antipsychotic medications — produces similar motor symptoms, referred to as drug-induced parkinsonism or extrapyramidal symptoms.
How does the nigrostriatal pathway relate to Parkinson’s disease?
Parkinson’s disease is fundamentally a disease of the nigrostriatal pathway. Its core pathology is the progressive degeneration of dopamine-producing neurons in the substantia nigra pars compacta, which progressively depletes striatal dopamine. The cardinal motor symptoms — bradykinesia, resting tremor, rigidity, and postural instability — are direct consequences of the resulting imbalance in the basal ganglia motor circuit. The effectiveness of levodopa (which replenishes striatal dopamine) and deep brain stimulation of the subthalamic nucleus (which compensates for the overactive indirect pathway) are both explained by their actions within this specific circuit. By the time motor symptoms become apparent, approximately 50–70% of SNc neurons have already been lost.
What is the difference between the nigrostriatal pathway and the mesolimbic pathway?
Both are dopaminergic pathways, but they serve distinct functions and originate in different brain regions. The nigrostriatal pathway originates in the substantia nigra pars compacta and projects to the dorsal striatum (caudate and putamen); its primary role is motor control. The mesolimbic pathway originates in the ventral tegmental area (VTA) and projects to the ventral striatum (nucleus accumbens), amygdala, and hippocampus; its primary roles are reward processing, motivation, and emotional learning. Disruption of the nigrostriatal pathway underlies Parkinson’s disease and drug-induced parkinsonism, while disruption of the mesolimbic pathway is implicated in addiction, schizophrenia, and depression. Many antipsychotic medications target both pathways simultaneously, which is why dopamine-blocking drugs reduce psychotic symptoms (via mesolimbic D2 blockade) but also impair motor function (via nigrostriatal D2 blockade).
What is the role of the striatum in the nigrostriatal pathway?
The striatum — composed of the caudate nucleus and putamen — functions as the primary input and integration hub of the basal ganglia and the principal destination of nigrostriatal dopamine. It receives excitatory input from virtually the entire cerebral cortex via the corticostriatal pathway, as well as dopaminergic input from the SNc. Striatal medium spiny neurons then process this convergent input through two populations: those expressing D1 receptors (participating in the direct pathway, which facilitates movement) and those expressing D2 receptors (participating in the indirect pathway, which suppresses movement). Dopamine modulates both populations simultaneously, amplifying direct pathway activity and dampening indirect pathway activity to produce a net pro-movement effect. The caudate is also involved in procedural learning and habit formation, while the putamen is more directly tied to real-time motor regulation.
Can the nigrostriatal pathway regenerate after damage?
Under natural conditions, the dopaminergic neurons of the substantia nigra pars compacta have very limited capacity for self-renewal or regeneration after damage — which is why Parkinson’s disease is progressive rather than self-limiting. This biological reality has been one of the primary drivers of research into neuroprotective and neurorestorative therapies. Neurotrophic factors, particularly GDNF (glial cell line-derived neurotrophic factor) and BDNF (brain-derived neurotrophic factor), have shown the capacity to support the survival of remaining SNc neurons in preclinical models, and several clinical trials have explored their delivery via surgical infusion or gene therapy vectors. Cell transplantation strategies — particularly those using iPSC-derived dopaminergic neurons — aim to replace lost neurons entirely. These approaches remain active areas of research and represent the most promising directions toward disease modification rather than just symptom management.
How do antipsychotic drugs affect the nigrostriatal pathway?
Antipsychotic medications — particularly first-generation (typical) antipsychotics like haloperidol — achieve their therapeutic effects by blocking D2 dopamine receptors, primarily in the mesolimbic pathway. However, D2 receptors are also abundantly expressed in the striatum, where they are essential for the indirect pathway’s normal function. When antipsychotics block these striatal D2 receptors, they produce a functional state similar to dopamine depletion in the nigrostriatal pathway, resulting in extrapyramidal symptoms (EPS): pseudoparkinsonism (tremor, rigidity, bradykinesia), akathisia, acute dystonia, and with prolonged use, tardive dyskinesia. Second-generation (atypical) antipsychotics were developed in part to reduce this nigrostriatal D2 blockade, achieving antipsychotic efficacy with lower EPS risk — though the tradeoff varies considerably across individual agents.
Bibliography
- Lanciego, J. L., Luquin, N., & Obeso, J. A. (2012). Functional neuroanatomy of the basal ganglia. Cold Spring Harbor Perspectives in Medicine, 2(12), a009621.
- Graybiel, A. M. (2008). Habits, rituals, and the evaluative brain. Annual Review of Neuroscience, 31, 359–387.
- Braak, H., Del Tredici, K., Rüb, U., de Vos, R. A., Jansen Steur, E. N., & Braak, E. (2003). Staging of brain pathology related to sporadic Parkinson’s disease. Neurobiology of Aging, 24(2), 197–211.
- Benabid, A. L., Chabardes, S., Mitrofanis, J., & Pollak, P. (2009). Deep brain stimulation of the subthalamic nucleus for the treatment of Parkinson’s disease. The Lancet Neurology, 8(1), 67–81.
- Purves, D., Augustine, G. J., Fitzpatrick, D., Hall, W. C., LaMantia, A. S., Mooney, R. D., Platt, M. L., & White, L. E. (Eds.). (2018). Neuroscience (6th ed.). Sinauer Associates.
- Albin, R. L., Young, A. B., & Penney, J. B. (1989). The functional anatomy of basal ganglia disorders. Trends in Neurosciences, 12(10), 366–375.
- DeLong, M. R. (1990). Primate models of movement disorders of basal ganglia origin. Trends in Neurosciences, 13(7), 281–285.
- Cotzias, G. C., Van Woert, M. H., & Schiffer, L. M. (1967). Aromatic amino acids and modification of parkinsonism. New England Journal of Medicine, 276(7), 374–379.
Use this citation format to reference the article clearly and help readers find the original source.
PsychologyFor. (2026). Nigrostriatal Pathway of the Brain: Structures and Functions. PsychologyFor. https://psychologyfor.com/nigrostriatal-pathway-of-the-brain-structures-and-functions/